Пищеварительная система ампуллярий приспособлена для питания водной растительностью. В первом приближении, она состоит из пяти отделов: захватывающий отдел (рот и ротовая полость с радулой и челюстями), область, где происходит предварительная обработка пищи (пищевод со слюнными железам, латеральный карман и зоб), пищеварительная область (трехкамерный желудок со связанными пищеварительными железами), область всасывания (кишечник) и изгнания (прямая кишка и анальное отверстие).
Рот ампуллярий, представляющий собой вертикальную щель, располагается между губными щупальцами и переходит в ротовую полость. Пища направляется губными щупальцами и, когда это необходимо, собирается ногой с поверхности воды.
Схема пищеварительной системы ампуллярий
Ротовая полость – мускулистая полость, усеянная кальцифицированными челюстями, с двумя наростами радулы и самой радулой (терка), расположена позади рта. Радула лежит на вершине наростов радулы (одонтофоры — хрящевая основа радулы) и покрыта несколькими рядами (26-53), каждый из которых включает 7 обновляемых хитиновых зубов. Когда одонтофоры раздвигаются, радула изгибается и растягивается, что приводит к рассредоточению зубов и трению. После того как пища попала в ротовую полость, мощные, известковые челюсти отгрызают кусок возле рта.
Челюсти (1), одонтофоры (2) и радула (3) Pomacea canaliculata.
Когда улитка ест, ротовая полость стягивается ко рту, и начинает функционировать весь сложный грызуще-скребущий механизм.
На иллюстрации можно видеть упрощенную модель движения рта ампуллярий, которая поможет понять принцип его работы. Стоит отметить, что радула улиток, в большей или меньшей степени, покрыта одонтофорами, которые изгибаются и растягивают её. Это обеспечивает движение зубов от- и навстречу друг другу, что позволяет соскребать пищу. Одонтофоры легко путаются с челюстями, которые сложно увидеть. Челюсти лишь слегка выглядывают изо рта, после чего радула задвигается и рот закрывается.
Последовательность движений: Рот открывается, челюсти нависают над радулой. Затем радула выдвигается вперед, а ряды зубов раздвигаются. Радула вращается подобно гусеничному траку на базе подстилающих одонтофоров. В определенной точке она выдвигается на максимальное расстояние, зубы стягиваются навстречу друг другу и трут пищу. Радула уходит обратно, тогда как челюсти выходят вперед и подобно ножницам отрезают кусочек корма. После этого, кусочек проходит в пищевод.
Радула ампуллярий имеет общую структуру, состоящую из ряда центральных или позвоночных зубов, боковых и двух пограничных рядов (внутренний и внешний ряды зубов) с каждой стороны радулы. Количество рядов, равно как и форма зубов, зависят от вида улиток [Martín S. M., Negrete L.H.L. 2007].
Радулы Pomacea canaliculata (1), Pomacea scalaris (2), Pomella megastoma (3) и Asolene platae (4) (илл. Martín S. M., Negrete L.H.L. 2007)
Благодаря богатым миоглобином мышцам, ротовая полость имеет красноватый оттенок. Миоглобин является дыхательным пигментом, который обеспечивает достаточное количество кислорода в метаболизме работающих мышц. Мышцы ротовой полости располагаются в передней части головного гемоцеля (большой сосудистый синус или полость).
Гемоцель — пространство между эктодермой и эндодермой, имеющееся у многих беспозвоночных и содержащее кровь (гемолимфу).
На тыльной стороне ротовой полости располагаются два латеральных кармана и проток слюнных желез. Светло-оранжевые слюнные железы лежат напротив широкого первого отдела пищевода (зоб) и продуцируют кислые мукополисахариды. Стенки зоба и пищевода покрыты продольными сгибами и содержат клетки, выделяющие нейтральные мукополисахариды и белки. Пищевод поворачивается на 180 градусов, соответствуя изгибу тела улитки, и соединяет ротовую полость с желудком. Пища перемещается по пищеводу за счет перистальтических движений.
Желудок ампуллярий представляет собой розовую U-образную структуру на левой стороне витка раковины. Стенка желудка состоит из 7 мышечных слоев, часто покрытых известковыми отложениями настилающих тканей или пищеварительных желез. Сама по себе, желудок включает три отдела: мышечный желудок (задняя камера — gizzard), преддверие (передняя камера — vestibule) и стебельчатый мешок (style sac).
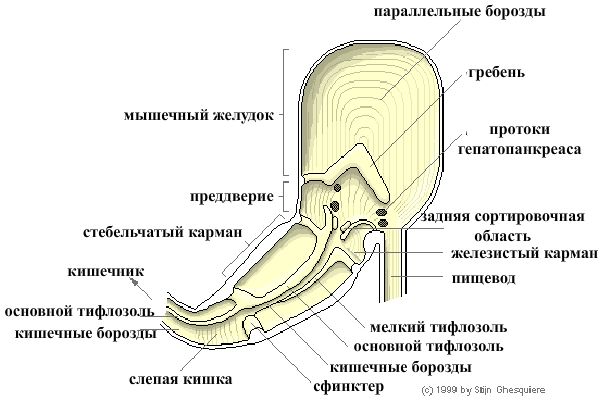
Пища смешивается с пищеварительными ферментами и перемалывается движениями мышц задней камеры желудка, соединенного с преддверием. Задний и передний отделы желудка отделены друг от друга толстым гребнем, который достигает крыши желудка, закрывая мышечный отдел. При сокращении мышц задней камеры, последняя объединяется с преддверием. Поверхность мышечного желудка имеет множество параллельных складок (борозд), а сама стенка включает 7 мышечных слоев.
Протоки пищеварительных желез или желез средней кишки, продуцирующих пищеварительные ферменты, открываются в ресничное преддверие. Железа средней кишки находится сверху желудка, и выровнена с семенником/яичником в верхних завитках раковины. Функции этой железы, известной как гепатопанкреас, аналогичны функциям печени (биохимические превращения питательных веществ и отходов) вместе с поджелудочной железой (образование пищеварительных ферментов) человека. Гепатопанкреас легко идентифицировать по темно-коричневому цвету.
У представителей родов Pomacea, Marisa и Asolene и, скорее всего, других ампуллярий, внутри этой железы присутствует большое количество зеленовато-черного вещества. При рассмотрении под микроскопом можно видеть два типа окрашенных элементов: клеточные округлые частицы (C corpuscles) с коричневато-зеленоватыми и темно-коричневыми похожими на туфельку элементами (K или kystic corpuscles). «C»-частицы имеют примерно 11-13 мкм в диаметре, а «K»-частицы – 35 мкм в длину и 13 мкм в ширину. Оба типа частиц также присутствуют не только в протоках и пищеварительном тракте, но также внутри железистых клеток, в кишечнике, экскрементах и даже просто в местах обитания улиток. В кишечнике «C»-частицы находятся в столбчатых эпителиоцитах, являющихся основной частью пищеварительных клеток, тогда как «K»-частицы связаны с экскреторными пирамидальными клетками. Вместе эти элементы составляют значительную часть железистой массы (11-13% от массы всех желез).
Частицы выделяются железистыми клетками в протоки и переносятся в желудок, где они смешиваются с пищей. В экскрементах частицы присутствуют, будучи смешанными с пищей, либо в составе плотных скоплений, практически на 100% состоящих из них. Помет в основном состоит из «C»-элементов. Фактически, эти элементы содержат ДНК улитки и двойную внешнюю мембрану, но недостаток ядра свидетельствует об их прокариотической природе.
Предполагаемое внешнее происхождение частиц в результате симбиоза исключается, потому что они обнаруживаются у улиток даже в стерильном хозяйстве. Кроме того, существование и развитие частиц внутри организма свидетельствует об их эндогенном происхождении. В частности, «C»-элементы образуются из сферических телец (вакуолей), которые, в свою очередь, развиваются внутри пищеварительных железистых клеток. Эти, лишенные мембраны, вакуоли выделяются в альвеолы и развиваются в зрелые «C»-элементы. В процессе прохождения кишечника они покрываются мембраной и трансформируются в «K»-частицы.
"C" и "K"-частицы из средней кишки Pomacea canaliculata
Внутри улитки находятся симбиотические прокариоты, существующие на взаимовыгодных условиях. Однако выгода для хозяина от их присутствия до сих пор неизвестна.
Железистый мешок преддверия содержит в своем эпителии множество мукопротеид-секретирующих клеток. В преддверии, спереди стебельчатого мешка (style sac). располагается сортировочный отдел, в котором отфильтровываются большие куски пищи. Вход в него окаймлен мощным мышечным сфинктером и соединяется с маленьким и большим тифлозолем, между которыми находятся глубокие, продольные кишечные борозды. Тифлозоли содержат реснички, образующие сильный ток по ходу кишечника, в то время как стенки стебельчатого мешка имеют вращающиеся поперек реснички, которые заставляют пищу двигаться по спирали.
Кишечник ампуллярии лежит близко к стебельчатому мешку, обвивается задней почкой и включает несколько отделов: слепая кишка, вторая и третья части кишечника и анальную папиллу.
Переваривание пищи частично происходит за счет ферментов, продуцируемой улиткой и симбиотическими микроорганизмами. Вещества всасываются через стенку кишечника, а затем переносятся с током крови. Задняя аорта обеспечивает кишечник кровью.
Между стебельчатым мешком желудка и слепой кишкой находится мышечный сфинктер, который контролирует перемещение пищи из желудка в кишечник. Ресничный основной тифлозоль начинается от преддверия желудка с кишечными бороздами, параллельными ему, и проходит через слепую кишку и первый отдел кишечника. Стенки первого отдела кишечника покрыты ресничными складками, ответственными за перемещение пищи. Кишечные борозды в данном отделе кишечника не имеют складок или ресничек. В конце первого отдела, они расширяются в мешок, тифлозоль сгибается в сторону мешка, а его носовая часть в сторону мелкого рубца второго отдела кишечника.
Мелкая не ресничная борозда второго отдела кишечника частично покрыт ресничными складками. Эпителий борозды включает железы, продуцирующие кислые мукополисахариды, что разительно отличает его от стенок других отделов кишечника, выделяющих нейтральные мукополисахариды.
Борозда, связанная с третьим отделом кишечника (прямая кишка) выпячиваются в небольшие дивертикулы (воронкообразное выпячивание стенки полого органа). Тифлозоль (гребень), окаймляющий борозду, становится очень широким, а сама борозда сужается и располагается на вершине гребня. Стенки третьего отдела кишечника имеют поперечные складки, лишенные ресничек. Близко к анальному отверстию в кишку открывается проток анальной железы, которая находится в основании анальной папиллы.
Анальная папилла расположена на крыше правой юбки мантии.
Рацион питания
Ампуллярии предпочитает мертвые, разлагающиеся растения, искусственные и живые корма. Они не поедают здоровые растения, кроме случаев, когда больше нет другой пищи. Это обстоятельство делает представителей семейства Ampullariidae полезным обитателем растительного аквариума.
Одно из исследований [Aditya G., Raut S.K. 2001] рациона питания распространенного аквариумного вида Pomacea bridgesi позволило четко определить пищевые предпочтения этой улитки. Интересно, что причиной проведения данной работы явилось внесение улиток из аквариумов в воды Индии и, в связи с этим, возникшая угроза конкуренции за пищевую нишу с местными видами.
В экспериментальных условиях готовились три аквариума (30х20х26 см), в которые на 23 см высоты наливалась вода. В каждый из них вносилось 30 улиток, 10 из которых принадлежали одной из трех групп: молодые (14-16 мм), половозрелые (30-32 см) и взрослые (38-40 мм). В течение недели ампуллярии питались латуком (Lactuca sativa), а затем начали получать корма различного типа и пропорций. Во всех случаях наблюдения продолжались 10 дней. Вода с фекалиями, пищевыми частицами, скелетами, костями и мертвыми улитками регулярно удалялись из аквариума с интервалами в 24 часа.
В независимости от групп, Pomacea bridgesi демонстрировали одинаковые пищевые предпочтения. Во всех случаях животная пища пользовалась более высоким спросом, чем растительная. Хотя трубочник (Tubifex sp.) и олигохеты (Branchiodrilus sp.) поглощались с одинаковой скоростью, живые черви были более предпочтительны, чем мертвые. Мертвые дождевые черви всегда потреблялись первыми в присутствии живых и мертвых Tubifex и Branchiodrilus. Полуразложившееся мясо креветок (десятиногие) и моллюсков, в независимости от вида, поедались в эквивалентных количествах, быстрее, чем олигохеты. В ряду тушек рыб пищевое предпочтение снижалось следующим образом: Setipinna phasa > Xenentodon cancila > Puntius ticto > Colisa fasciatus > Ambylopharyngodon mola > Labeo rohita > Labeo bata. В независимости от вида, рыба поедалась быстрее, чем моллюски и креветки. Наконец, ампулирии демонстрировали избирательность при поглощении растений: L. sativa > Solanum tuberosa > Rumex vascicarius > Brassica juncea > Raphanus sativus. Виды Cobamba sp. и Eichhornia crassipes потреблялись в эквивалентных количествах.
Результаты продемонстрировали широкий диапазон кормовой базы улиток Pomacea bridgesi, которые без проблем приспособились и образовали колонии в водах Индии. В результате, макрофитофаги, зоофаги и микрофаги столкнулись с серьезной конкуренцией за пищу.
——
www.applesnail.net/content/anatomy/digestion.php
Aditya G., Raut S.K. Food of the snail, Pomacea bridgesi, introduced in India. Current Science 80 : 919–21. 2001.
Martín S. M., Negrete L.H.L. Radular ultrastructure of South American Ampullariidae (Gastropoda: Prosobranchia). Braz. J. Biol. vol.67 no.4 São Carlos Nov. 2007.

Реакция постоянных читателей: