Забота за потомством является сложной задачей, особенно если вы являетесь самкой цихлид.
Традиционная для кратерного озера в Никарагуа, цихлида заботится о своих мальках, защищая их от хищников. Ученые показали, что самец и самка цихлиды не всегда разделяют родительские заботы.
Новое исследование, проводимое международной командой, включающей два биолога из университета Монаша, и опубликованное в журнале «Behavioral Ecology and Sociobiology», показало, что самцы цихлид имеют склонность покидать своих партнеров, которые в одиночку осуществляют заботу о потомстве.
В попытках понять экологические факторы, определяющие это поведение, ведущий автор, доктор Топи Лихтонин (Topi Lehtonen) из университета Турку в Финляндии, провел много часов под водой, наблюдая родительское поведение различных цихлид.
Доктор Лихтонин и его коллеги, включающие доктора Боба Вонга (Bob Wong) и доктора Андреаса Свенссона (Andreas Svensson) из университета Монаша, а также профессора Аксела Миера из университета Констанц в Германии, обнаружил, что отказ от выполнения родительских обязательств у партнеров более выражен среди крупных видов цихлид, населяющих озеро, и феномен имеет место в сезон размножения.
Доктор Боб Вонг, старший лектор на научном факультете при университете Монаша назвал причины, из-за которых самец покидает своих партнеров и мальков, что было не совсем ясно. Вероятно, подобное поведение связано с затратами и выгодами между полноценной заботой о потомстве и доступностью дополнительных возможностей спаривания.
«Не смотря на мотивы, разрыв пар производителей является плохой новостью для покинутой самки и её мальков, » сказал доктор Вонг.
Исследование обнаружило, что уход самцов точно определяется тем, насколько самка способна выполнять свои обязательства, в частности, по защите своих мальков от крупных хищников, таких как большеротый гобиоморус.
«Находка вносит значительный вклад в механизм системы воспроизводства в животном мире, эволюции родительской заботы, затратами и выгодой от покидания мальков,» сказал доктор Вонг.
——
источник: sciencedaily.com/releases/2010/09/100920101151.htm
Чуть позже появится перевод исследования, озвученного выше.
Добавлено 23.10.2010
Возможно, в некоторых местах перевод хромает, но вы можете ознакомиться с оригиналом статьи, которая представлена в интернете в свободнном доступе.
Приспособление по заботе о потомстве в отсутствии самца у двух видов цихлид из кратерного озера в Никарагуа.
Во многих таксонах родительские стратегии могут различаться среди особей. Это особенно характерно для видов с заботой обоих родителей, с самцами, которые чаще, чем самки покидает своих партнеров. Не смотря на изобилие теоретических прогнозов и эмпирических данных на факторы, провоцирующие самца на оставление самок, много меньше известно о следствие, которое отражается в поведении самки, в частности, в полевых и нептичьих системах. Здесь, сравнивается уровень защиты потомства, типы защиты потомства и успешность выводка при одиночной или парной заботе неотропических цихлид в их естественной среде обитания. В отношении к степени территориальной агрессии против потенциальных хищников, самка способна полностью компенсировать отсутствие самца, но при этом она охраняет более миниатюрную площадь, которая является адекватной потомству, по крайней мере, двух видов. Результаты работы свидетельствуют о том, что даже большая пространственная компенсация за счет родительской заботы одной самки не может быть достаточна для обеспечения адекватной качественной компенсации при недостатке самца, выделяя значимость разграничения между двумя данными аспектами родительской опеки.
Введение
Моногамное поведение часто связано с заботой обоих родителей за потомством (Smith andWootton 1995; Reynolds 1996). Согласно этому, условия окружающей среды и социальные факторы, которые приводят к увеличению относительной выгоды от полигинии, могут также провоцировать оставление самцом своих отпрысков (Smith and Wootton 1995; Gross 2005). Факторы окружающей среды, влияющие на возможность сохранения потомства, могут также определять, сможет ли самка, будучи покинутой, длительное время заботиться о мальках (Lazarus 1990). Факты, однако, говорят о том, что покинутая самка часто остается со своими мальками (McNamara et al. 2002). Более того, у большинства видов с заботой обоих родителей уход самца документально отмечался намного чаще, чем самки (для примера виды, у которых любой пол с равной вероятностью может покинуть. см. Bessinger and Snyder 1987; Griggio and Pilastro 2007).
Результаты полового конфликта в области заботы родителей притягивает внимание биологов (e.g. Clutton-Brock and Vincent 1991; Queller 1997; McNamara et al. 1999; Kokko and Jennions 2008). Метод, воспроизведенный в работе, экспериментально снижает уровень участия самца в родительской заботе, а затем проводит мониторинг реакций самок, находящихся под контролем лабораторных условий (Houston et al. 2005). Тем не менее, полевые исследования являются ключевыми, если мы желаем понять, как одинокие самки могут компенсировать отсутствие самца при экологических факторах. В одном из таких исследований, самки Испанского воробья (Passer hispaniolensis) прикладывают огромные усилия в обеспечение птенцов, сопоставимое с парой самок, но она не в состоянии выполнять работу в той же степени, в какой её выполняют самки вместе с самцами (Marques 2004). Обобщая, многое из того, что мы знаем о компенсации родительской заботы почти всегда касается информации, добытой при изучении птиц (Harrison et al. 2009). Иными словами, наши представления о данном феномене в других таксонах изучены в меньшей степени.
Более того, предшествующие исследования покидания самцами самок обычно сосредоточены не только на изменениях в количественных характеристиках заботы, в то время как видится возможность рассмотрения эффектов на качестве предоставляемой заботы. Например, при изучении птиц обычно измеряется только режим кормления без качественных характеристик, либо просто количество пищи, приносимое в гнездо (Harrison et al. 2009). Последнее может быть важно, если, например, имеется компромиссное соотношение между количеством и качеством компонентов родительской компенсации. Отсюда, один не принимается, априори, позитивная корреляция между данными двумя аспектами заботы (т.е. количество против качества). Более определенно, оценивается только один, в то время как игнорируется другой, что не дает нам понять результаты оставления потомства самцом. Согласно этому, в данном полевом исследовании использовались «нептичьи» системы изучения для рассмотрения того, насколько способна расширить свои обязанности одинокая самка при защите потомства после того, как самец долгое время не появлялся, и выделить качественные и количественные аспекты компенсаторной заботы.
Методы
Полевые исследования проводились на двух видах цихлид эндемиков кратерного озера в Никарагуа, Центральной Америки: Amphilophus sagittae из озера Ксилоа (Stauffer and McKaye 2002; Elmer et al. 2009) и Amphilophus zaliosus (стрельчатая цихлида) из озера Апойо (Barlow and Munsey 1976; Barluenga et al. 2006). Несколько особенностей биологии данных видов делает их особенно интересными в плане изучения различий в родительской заботе одинокой самки и находящейся в паре. Оба вида принадлежат к комплексу Мидас цихлид (Elmer et al. 2010), члены которых ранее рассматривались как моногамные с заботой обоих родителей (McKaye 1977; Rogers 1987, 1988; Barlow 2000), однако недавно в природных условиях было обнаружена также вовлеченность в заботу над потомством лишь самок (Lehtonen et al. 2010b). Подобно другим членам данного комплекса видов, A. sagittae и A. Zaliosus имеют продолжительный сезон размножения и установленные постоянные территории для его проведения (Barlow 1976; McKaye 1977; Rogers 1987; Barlow 2000), что делает их крайне легким объектом для полевых наблюдений. Свободно плавающие мальки данной рыбы являются достаточно независимыми для самостоятельного питания планктоном (Barlow 1976) и, иногда, слизистым выделением эпителия родителей (Noakes and Barlow 1973; персональные наблюдения). Отпрыски, однако, полагаются на своих родителей в защите против конспецифичных и гетероспецифичных хищников (Barlow 1976; Rogers 1987; McKaye and Murry 2008; Lehtonen et al. 2010a), а успешность обороны является критичным критерием адекватности родительской заботы (McKaye 1977; Rogers 1987, 1988; Barlow 2000). В этом отношении, как количество (т.е. степень агрессивных ответов), таки качество опеки (т.е. дистанция реакции и типы агрессивного поведения, см.ниже), вероятно, оба являются важными.
Все наблюдения проводились между декабрём 2007 и январем 2008 года на глубинах 3,5-18 метров, с использованием SCUBA. Обратившись к вопросу ответов самки при оставлении её самцом, исследователи сравнили весь объем работы и другие аспекты поведения по защите потомства одинокими и самками, находящимися в паре, для A. sagittae (одинокие=5, парные=21) и стрельчатой цихлиды (одинокие =14, парные =15), и изучили возрастной разбег потомства от самок озвученных типов (т.е. одиноких против парных). Пропорции одиноких и парных самок включили в работу, отражающую частоту, с которой они наблюдались в поле (см. Lehtonen et al. 2010b). Родительское поведение оценивалось в соответствии со всеми, а не одним выводком стрельчатой цихлиды, описываемым по ходу исследования. Мальки A. Sagittae выбраны случайно в подгруппе изучаемого потомства, которое было не старше 3 недель (примерный возраст) у двух видов. Так как вероятность покидания самцом потомства возрастает с увеличением оберегающих факторов (Smith and Wootton 1995; Gross 2005), это заставляет ожидать, что одинокие самки, оберегающие потомство будут в среднем крупнее и старше. Тем не менее, если количество мальков, защищаемых одинокой самкой, является явно меньшим, чем у парных особей, можно ожидать противоположную тенденцию.
Были приспособлены простейшие протоколы, которые успешно использовались на ранней стадии исследования (Lehtonen et al. 2010a). В частности, после локализации территории размножения, представляющая собой область площадью примерно 2 метра, забота не прекращалась родителями или любыми соседними размножающимися цихлидами. Записывались данные, такие как, глубина воды и статус самки (одинокая, парная). Для большинства территорий (Таблица 1) также приблизительно отмечалась общая длина самок, и возраст свободноплавающих мальков (начиная с «0» лет для ювенильных, которые только начали передвигаться). Этот возраст оценивается на основе видимого размера мальков и калибровался против предыдущих наблюдений стадий развития, как в природе, так и в лаборатории. Кроме того, записывалась активность производителей в течение 15 минут после акклиматизационного периода, который составляет минимум 3 минуты. Каждый акт поведения, проявляющегося в защите территории классифицировался согласно одному из следующих трёх категорий: «демонстрация» (растопыренные плавники и жаберные крышки без- или с постепенным движением навстречу вторгшейся рыбке), «атака» (стремительное движение к нарушителю и преследование до полного изгнания) и «преследование» (атаки, которые продолжаются даже, когда нарушитель спасается бегством). Данные поведенческие ответы рассматривались в отношении от их различий эффективности и стоимости. Так, демонстрация, имеющая низкий уровень затрат энергии и столкновения, и преследование, имеющее высокий уровень данных показателей. Для каждого из трех актов агрессии была отмечена дистанционная реакция, измеряемая от центра территории, в качестве примерной аппроксимации ревира. Для демонстрации и атак, эта характеристика оценивалась как дистанция между (приближенно) центра выводка до локализации рыбки нарушителя ко времени ответа. Для преследования были приближенно вычислены дистанция между центром ревира и производителем, находящегося в конечной точке преследования.

Таблица 1. Длина самок и возраст выводка у одиноких особей и находящихся в паре. Значения представлены как среднее ± стандартная ошибка.
Используя выше представленные данные, появилась возможность оценить (1) общую степень (количество) защиты мальков (анализировалась как сумма всех демонстраций, атак и преследований, имеющих место быть в 15-минутный период наблюдения), (2) качество защиты мальков по продолжительности дистанционной реакции и относительным частотам трех категорий агрессии (демонстрация, атака и преследование), и (3) успешность самки в защите потомства, как «возрастной счёт», предоставляемый каждому выводку (использовались данные от всего множества выводков за весь сезон размножения). Конечный уровень защиты потомства анализировался с использованием линейных моделей видов (Стрельчатая цихлида или A. sagittae), статуса (одиночная или парная) и парного взаимодействия данных факторов как предсказателя изменчивости. Подсчитанные данные возводились в квадратный корень для удовлетворения нормальности, и применялся t-тест для сравнения тестовых параметров. Кроме того, анализировалось, может ли различаться итоговый уровень у одиночной и парной самки (позднее исследование, основанное на попытке коллективно наблюдать как за самками, так и за самцами в паре). Для изучения того, может ли тип агрессии отличаться у одиночной и парной самки, анализировались данные трех типов агрессии в линейной смешанной модели. Данная модель подходит для максимизации логарифмического правдоподобия, с видами, статусом и категорией агрессии в качестве констант, и идентификацией территории в качестве переменного фактора. Незначимые сроки удалялись следующей моделью упрощения, использующей критерий отношения правдоподобия (G2 с α=0,05, Crawley 2007). Затем проводилась проверка того, отличаются ли относительные частоты трех категорий агрессии у одиночной особи и самки, находящейся в паре с самцом. Данное тестирование осуществлялось при помощи анализа расчетов всех трех типов между парами форм (т.е. демонстрация против атак, демонстрация против преследования и атаки против преследования), с использованием обобщенных смешанных моделей с биноминальной ошибкой и функцией, связанной с логит-преобразованием. Каждое сравнение имело биноминальную частоту от двух типов агрессии в качестве зависимой переменной, статуса и видов в качестве констант, и территории в качестве переменного фактора. Были представлены три модели, каждой из которых, применялся один или три возможных сравнения (демонстрация против атак, демонстрация против преследования и атаки против преследования). Данная процедура эквивалентна выполнению полиномного (триномиального) анализа, однако проще в истолковании. Модельное упрощение проводилось с использованием χ2 тестов (Crawley 2007). Наконец, для оценки различий в дистанции реагирования (доверенный размер территории), длины самок и возраста мальков (логарифм преобразовывался) между одиночной особью и самки, находящейся в паре с самцом, выполнялся анализ вариации (ANOVA), изначально используя данные наблюдений в качестве ковариации. Тем не менее, так как эффект данных наблюдений во всех случаях оказывался незначимым, позднее он был исключен из анализа. Тест ANOVA производился в программе Systat 12 (SPSS Inc.), а для сбора анализов использовалась ПО R 2.8.0 (R Development Core Team).
Результаты
Уровень защиты потомства (количество заботы)

Таблица 2. Результаты двухфакторного анализа ANOVA. Эффект «Статуса» связан с тем, была ли самка одинокой или находилась в паре.
Одинокая и парная самки обоих видов не имели значимых различий в размере тела (Таблица 1 и 2). Вследствие этого, размеры не были включены в качестве ковариации в любой из проверок. Не имелось значимых связей между видами и статусом в общей массе (через 15 минут) агрессивного поведения (F1,51=0,362, p=0,55). Модель, построенная без взаимосвязей, продемонстрировала значительный эффект статуса (t52=3,65, p<0,001), но не видами (t52=0,512, p=0,61) в общем уровне агрессии. Другими словами, одинокие самки проявляли повышенный уровень агрессивного поведения при защите потомства по сравнению с парными самками обоих видов (Рисунок 1). Когда в анализ включался вклад самцов, не имелось большого эффекта статуса на уровень агрессии (t52=1,28, p=0,21). Это значит, что одинокая самка полностью компенсировала недостаток участия самца в агрессивном поведении, и её уровень был схож с общим уровнем агрессии, производимым парой производителей (Рисунок 1).
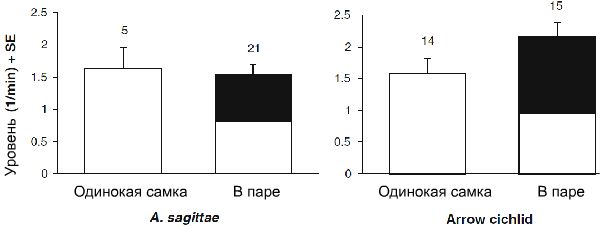
Рисунок 1. Итоговый уровень агрессивных ответов по отношению к потенциальным хищникам. Белые столбцы показывают итоговый уровень агрессии самок (как родиноких, так и в паре), а черным указано участие самца. Столбцы ошибок «В паре» складывается для обоих полов. Размер выборки указан числом над каждым столбцом.
Качество защиты потомства
Что касается трех критериев агрессивности (т.е. демонстрация, атаки и преследование), не имелось значимых трехсторонних взаимосвязей между видами, статусом и агрессивной категорией (G2=1,98, p=0,37), и не имелось значимой взаимосвязи между видами и статусом (G2=0,467, p=0,49). Тем не менее, имелись значимые связи между категорией агрессии и статусом (G2=19,0, p<0,001), а также между категорией агрессии и видами (G2=18,2, p<0,001). Другими словами, частота трех категорий агрессии различалась между одинокими и парными самками, также как и между двумя видами.
Для рассмотрения того, что могло производить данные взаимосвязи, исследовались относительные частоты трех типов агрессии с использованием биномиальных смешанных моделей. Результаты моделей не показали значимых взаимосвязей между статусом и видами (все χ2<3,52, p>0,05), и, таким образом, они были преобразованы без данных элементов. Редуцированные модели показали, что по сравнению с одинокими самками, парные особи выполняют больше преследований, чем демонстраций (z=2,63, p=0,009) и больше преследований, чем атак (z=2,51, p=0,012, Таблица 3). Тем не менее, количество демонстраций по отношению к атакам не зависело от статуса самки (z=0.505, p=0.61). Два вида не отличались по относительным частотам преследований к демонстрациям (z=1,25, p=0,21) или преследований к атакам (z=1.62, p=0,11). Однако, стрельчатая цихлида производила больше атак относительно демонстраций по сравнению с A. Sagittae, независимо от статуса самки (виды: z=4,43, p<0,001; Таблица 3).
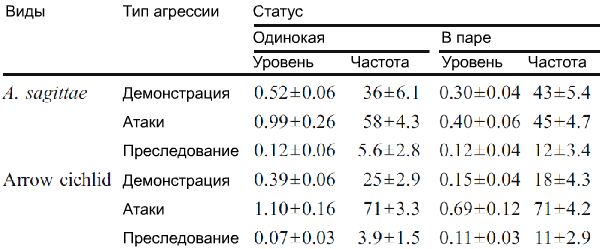
Таблица 3. Распределение трех категорий агрессии у одиноких и парных самок, представлено как уровень (событие/мин), так и в относительной частоте (%). Выборка для одинокой самки и самки в паре составляла 5 и 21 для A. sagittae, и 14 и 15 для стрельчатой цихлиды, соответственно. Все значения представлены как среднее ± стандартная ошибка средней.
Дистанция реагирования к соответствующим хищникам отличалась незначительно у одиноких и парных самок для обоих видов (Рисунок 2; Таблица 2).
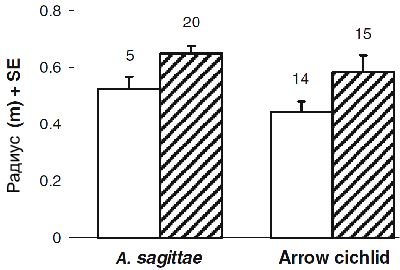
Рисунок 2. Радиус ревира, охраняемого самкой. Белые столбцы относятся к одиноким самкам, заштрихованные — к парным. Размер выборки представлен числами.
Успешность защиты потомства
Для обоих рассмотренных видов, возраст мальков, охраняемых как одинокой, так и парной самкой не различался (Таблица 1 и 2). Тем не менее, в случае A. sagittae, на протяжении всего сезона размножения, охраняемые мальки одиноких самок были мельче, чем у самок, находящихся в паре с самцом (Таблица 1 и 2; post hoc: t23=2,21, p=0,038).
Обсуждение
Ранние исследования, в большинстве своем на птицах, показали, что самки, будучи загруженными своими обязанностями, при снижении участия самца в заботе над потомством, способны (или стремятся) лишь частично компенсировать отсутствие партнера (Houston et al. 2005; Harrison et al. 2009). Напротив, данное исследование продемонстрировало, что одинокие самки цихлид имеют возможность полностью компенсировать отсутствие самцов в отношении степени агрессивных ответов к потенциальным хищникам. В частности, было обнаружено, что одинокие самки достигают примерно такого же уровня ответов, которые наблюдаются у пары производителей (т.е. основанное на коллективной деятельности как самца, так и самки в паре). Тем не менее, несмотря на эту активную количественную компенсацию в уровне работы, одинокие самки не способны обеспечить такую же степень качества заботы, которая проводится парой производителей. Так, обнаружено, что одна самка у обоих видов показывала укороченную дистанцию реагирования и использовала менее агрессивные/затратные оборонительные реакции (низкий процент преследований) по сравнению с парными рыбками. Оборона меньшей территории или невозможность оставить потомство при преследовании хищников, что представляется главной опасностью для мальков, может быть препятствием в качестве защиты. Отсюда, результаты подчеркивают важность рассмотрения того, может ли самка компенсировать как количество, так и качество заботы, которое в большинстве случаев игнорировалось в исследовательских работах, и на что было обращено особое внимание (Harrison et al. 2009).
Согласно измерениям в сезон размножения, у A. Sagittae средний размер мальков, охраняемых одинокими самками, был меньше, чем у мальков, поддерживаемых парой производителей. Потомство одиноких особей могло просто не обладать такой выживаемостью как мальки пары родителей в случае, когда самка в одиночку пыталась обороняться от хищников. Тем не менее, нельзя вычеркивать возможность того, что мальки одиноких самок могли также развиваться, но вследствие нехватки корма, меньшими темпами. Даже если цихлиды напрямую не участвуют в кормлении своих мальков, одинокие особи A. Saggitae были менее результативны в отношении количества корма из-за меньшего охвата территории или обеспечения достаточного количества слизи для питания своего потомства.
В противоположность подсчетам для A. saggitae, не было обнаружено различий в среднем размере выводка у одинокой и парной самок стрельчатой цихлиды (A. zaliosus). Предполагается, что это вызвано межвидовыми различиями в живучести потомства парных особей. Во время исследования (сезон размножения 2007-2008 год) выводок стрельчатой цихлиды имел очень низкий уровень выживаемости (независимо от статуса самки), в то время как потомство A. Sagittae обладало более высокой выживаемостью (Lehtonen, McCrary и Meyer, неопубликованная рукопись). Более того, полевые исследования, проведенные на других Неотропических цихлидах, показали, что самцы, в основном, не покидают потомство вплоть до момента, когда мальки переходят к свободному плаванию (Wisenden 1994; Jennions и Polakow 2001; Vélez et al. 2002; персональные наблюдения в Никарагуа). Исходя из этого, не ожидалось различий в выживаемости выводка между двумя категориями самок до этапа, когда мальки в течение нескольких дней полностью переходят к свободному плаванию.
Остается под вопросом, почему одинокие самки цихлид проявляют повышенную количественную компенсацию в условиях отсутствия самца, чем самки большинство других животных, изученных в ряде работ (Houston et al. 2005; Smiseth et al. 2005; Harrison et al. 2009; Suzuki and Nagano 2009). Одной из возможных причин является то, что предыдущие исследования были сосредоточены на инкубации выводка и обеспечении пропитания, в то время как, если одинокая самка цихлид не компенсирует уровень заботы во всех направлениях, то потомство будет иметь чрезвычайно низкую выживаемость, благодаря хищникам и высокой конкуренции на территории (McKaye 1977; Rogers 1988). Более того, взаимосвязь между числом отпрысков и затратами на родительскую заботу, вероятно, различна у рыбок по сравнению с птицами, либо, в целом, по сравнению со многими другими группами животных (Lazarus и Inglis 1986; Wisenden 1999; Rauter и Moore 2004). Тем не менее, множество работ, предпочтительно на обширном диапазоне таксонов, нуждаются в оценке того, как динамика стоимость/эффективность опеки влияет на количественные аспекты компенсации, относящиеся к одиноким самкам.
Так как бесконтактный метод данного изучения не позволяет экспериментально исключить самцов по этическим и практическим причинам, он видится пригодным, когда необходимо одиноких самок, включенных в исследование, представлялись в качестве неслучайной подгруппы самок в популяции. Однако, ученым представляется невероятным, что «качество» одиноких особей является очень предвзятым, либо, на самом деле, любая подобная предубежденность может объяснить наблюдаемые паттерны в защите потомства. Во-первых, были обнаружены незначимые отличия в размере тела у одиноких и парных самок. Это важно потому, что размер тела, как известно, коррелирует с измерениями качества у Неотропических цихлид, особенно в отношении количества икринок и родительской заботы (McKaye 1986; Wisenden 1995; Barlow 2000). Во-вторых, вопреки ожиданиям исследователей, если одинокие и парные самки различались в качестве, изменение в поведении модели соответственно не указывает на нижнюю планку (короткая дистанция реагирования, низкая активность преследования) или верхнюю (высокая степень агрессивных ответов), нежели среднее качество. Отсюда, результаты свидетельствуют о модификации поведения одиноких самок для того, чтобы выносить нагрузки при защите потомства.
В заключении, результаты исследования показывают, что когда самки остаются для заботы за потомством, даже полная компенсация уровня родительской работы деятельности неравна по качеству заботы парной особи. Результаты говорят о том, что необходимо рассматривать не только количественные, но и качественные аспекты компенсации опеки.
——
springerlink.com/index/X6041W587627HK16.pdf
Похожие статьи: