
Чешуя является важным хранилищем кальция и фосфора и наряду с кожей выполняет функцию барьера от окружающей среды и повреждающих факторов. Живой неорогевающий эпидермис и чешуя являются функционально специализированной костной кожей, образующей дермальные скелетные элементы после метаморфоза молоди рыб. Стоит отметить, что ранние этапы развития покровов у водных и сухопутных позвоночных животных схожи [5].
У костных рыб кожа представлена тремя слоями: эпидермис, дерма и подкожная клетчатка, толщина которой очень вариабельна. Чешуйки обособлены и находятся в кармашках, которые, в свою очередь, располагаются в поверхностной дерме, направлены вглубь её и выстланы тонким слоем эпидермиса.
Чешуя большинства костных рыб классифицируется как эласмоидная и состоит из тонкого наружного, хорошо кальцинированного, так называемого, костного слоя, и толстой внутренней, частично кальцинированной, базальной пластинки. Структура костного слоя напоминает сотканную кость; она состоит из кристаллов гидроксиапатита и случайно ориентированных коллагеновых волокон типа I с обильным количеством основного вещества. Базальная пластинка состоит из множества тонких ламелл, каждая из которых включает плотноупакованные толстые коллагеновые волокна типа
I постоянного диаметра и небольшое количество основного аморфного матрикса. Кроме того, она покрыта эласмобластами (формирующие чешую клетки) и остеокластами (рассасывающие чешую клетки). Характерной особенностью базальной пластинки является трехмерное распределение её коллагеновых волокон. Они параллельны в пределах одной ламеллы, тогда как между собой ламеллы имеют разно ориентированные волокна. У некоторых видов пластинка стала ортогональной, похожей на клееную фанеру.
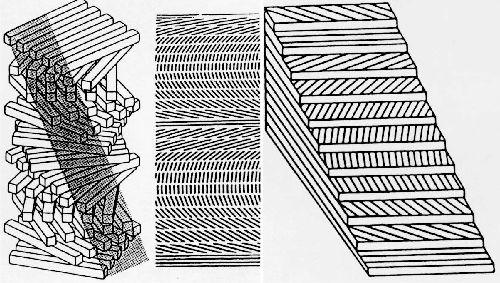
«Фанеро»-подобная ориентация ламелл (иллюстрация — www.engin.umich.edu/class/bme456/bonestructure/bonestructure.htm)
Характерное «фанеро»-подобное выравнивание коллагеновых волокон в базальном слое чешуи также наблюдается в строме роговицы. Более того, волокна данных структур обладают на удивление одинаковым диаметром. Уникальное свойство, прозрачность роговицы, достигается наличием двух особенностей волокон стромы — их одинаковым диаметром и специфическим выравниванием. Очевидным отличием роговицы глаза от базальной пластинки является поздняя кальцинация, происходящая у последней. Предполагается, что регенерация базального слоя чешуи может стать моделью процессов происходящих при восстановлении роговицы глаза [4].
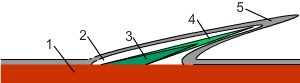
На иллюстрации: 1. Дерма; 2. Чешуйный карман; 3. Базальная пластинка чешуи; 4. Костный слой чешуи; 5. Эктодерма.
При потере чешуи рыбка утрачивает часть эпидермальных клеток, саму чешую и поверхностный слой дермы. Данное повреждение быстро заживляется, кожа покрывается слизью, а от границ раны начинается процесс восстановления эпителия, заканчивающийся в течение нескольких часов [3]. Кроме того, спустя несколько недель полностью восстанавливается чешуйчатый покров [4; 2]. Стоит отметить, что скорость восстановления поврежденной чешуи гораздо выше, чем её нормальное развитие в онтогенезе.
Процесс регенерации подразделяется на 4 этапа: образование нового эпителия и дифференцировка формирующих чешую клеток (1-2 дня), быстрая продукция матрикса внешнего (костного) слоя (3-5 день), продукция матрикса базальной пластинки (6-14 день) и, наконец, частичная минерализация базальной пластинки (проходит очень интенсивно, 14-28 день) [4].
Например, у золотой рыбки (Carassius auratus) наиболее быстрое восстановление чешуи наблюдается в течение первых 5 дней после повреждения. Постепенное снижение темпа прироста площади чешуйки связано с постоянным линейным увеличением её массы за период 5-28 дней. Приоритет от увеличения площади постепенно смещается в сторону присоединения базальной пластинки. В процессе восстановления, отложение кальция и фосфора происходит практически линейно. Кальцинация костного слоя предшествует кальцинации базального слоя, которая начинается спустя 14 дней после регенерации. Кальцинация базальной пластинки начинается и постепенно нарастает. Через 28 дней концентрация кальция и фосфора в новых чешуйках составляет 72% от онтогенетической чешуи. Позднее начало восстановления и медленная скорость кальцинации базальной пластинки определяет низкую скорость восстановления состава кальция и фосфора [4].
Не смотря на то, что гистология регенерации чешуи костных рыб активно исследовалась у нескольких видов карповых Cyprinidae, карпозубых (Cyprinodontidae) и цихлид (Cichlidae), в литературе плохо освещены тонкие клеточные механизмы восстановления повреждения. Одна из работ, в которой определялся состав пролиферирующих клеток и их вовлеченность в регенерацию, была проведена с двухточечным хромисом (Hemichromis bimaculatus) в качестве объекта исследования. Весь процесс восстановления чешуйчатого покрова отслеживался при помощи сканирующего электронного микроскопа. Было обнаружено, что первыми рану окружают склеробласты, популяция которых позднее заполняет пустые чешуйные карманы. Вначале формируются удлиненные сосочки, вероятно, из дифференцирующихся дермальных фибробластов. В середине сосочка появляется и становится пульпой зачаток чешуйки костного слоя [1].
По мере развития и накопления коллагеновых волокон костная пластинка приобретает округлую форму, по её краям можно обнаружить склеробласты. Верхняя часть костного слоя в дальнейшем обызвествляется и становится твердой. Базальная пластинка поздно кальцинируется и вследствие этого сохраняет волокнистую структуру, что позволяет поступать новым клеткам, которые формируют чешую
[6].
Все окружающие крупные склеробласты вовлечены в формирование чешуи, хотя позднее три популяции специализируются согласно своему местонахождению вокруг неё. Наружные склеробласты уплощаются, когда костный слой достигает своей окончательной толщины; глубокие — отвечают за формирование базальной пластинки, в то время как пограничные склеробласты увеличивают диаметр костного слоя.
В процессе регенерации склеробласты более многочисленны, чем во время отногенетического развития чешуи. В частности, глубокие склеробласты продуцируют столбчатый эпителий при основании базальной пластинки, однако они не наблюдаются в онтогенезе. Кроме того, регенерирующая базальная пластинка обладает ортогональной «фанеро» — подобной структурой, что никогда не обнаруживается в эмбриональном развитии, где данный тип структуры является промежуточным.
Формирование матрикса поврежденной чешуи отличается от его нормального развития в онтогенезе, что связано с продуцирующими волокна склеробластами, которые выглядят призматическими при формировании и становятся плоскими на поздней стадии регенерации [1].
Примечание:
Термины, используемые для обозначения клеток, которые участвуют в образовании эласмоидной чешуи.
1. Фибробласт (фиброцит) продуцирует межклеточный матрикс в системе тканей внутренней среды;
2. Склеробласт является клеткой фибробластического ряда, участвующие в образовании костных пластинок чешуи рыб;
3. Остеобласт (остеоцит) является клеткой костной ткани;
4. Эласмобласт формирует эласмридную чешую.
——
1. Jean-Yves Sire, Jacqueline Géraudie. Fine structure of regenerating scales and their associated cells in the cichlid Hemichromis bimaculatus (Gill). Cell and Tissue Research, 237 (3) : 537-547. 1984.
2.
Bereiter-Hahn J, Zylberberg L: Regeneration of teleost fish scale. Comparative biochemistry and physiology. 105A : 625-641. 1993.
3. Iger Y, Abraham M: The process of skin healing in experimentally wounded carp. Journal of Fish Biology. 36 : 421-437. 1990.
4. Ohira Y, Shimizu M, Ura K, Takagi Y. Scale regeneration and calcification in goldfish Carassius auratus: quantitative and morphological processes. Fisheries Science. 73 (1)
: 46-54. 2007.
5. Campinho M.A, Silva N, Sweeney G.E, Power D.M. Molecular, cellular and histological changes in skin from a larval to an adult phenotype during bony fish metamorphosis. Cell Tissue Res. 327
: 267-284. 2007.
6. Сидорова Е. И. Морфологическое и авторадиографическое исследование регенерирующей чешуи у Carassius auratus. 2004
Строение кожи Sparus auratus — www.biomedcentral.com/1471-2164/12/490
Похожие статьи:
Восстановление поврежденных структур Фундулуса гетероклитуса