 Каким образом паразиты находят и определяют хозяина хорошо известно для церкарий трематод. Данные многоклеточные организмы отвечают комплексом поведенческих реакций на последовательности различных химических сигналов хозяина, хотя при этом происходит множество этапов внедрения. Тем не менее, мало известно о поисковом поведении одноклеточных паразитов. Руководствуясь потребностью в рассмотрении данного вопроса, была проведена работа, результаты которой опубликованы в 1998 году в «Токайском журнале экспериментальной и клинической медицины» (The Tokai journal of Experimental and Clinical Medicine) в статье «Theronts of Ichthyophthirius multifiliis find their Fish Hosts with Complex Behavior Patterns and in Response to Different Chemical Signals». Авторы изучили особенности поисковых реакций инвазивных теронтов I. multifiliis во время представления им различных фракций и химических модификаций кожи рыб.
Каким образом паразиты находят и определяют хозяина хорошо известно для церкарий трематод. Данные многоклеточные организмы отвечают комплексом поведенческих реакций на последовательности различных химических сигналов хозяина, хотя при этом происходит множество этапов внедрения. Тем не менее, мало известно о поисковом поведении одноклеточных паразитов. Руководствуясь потребностью в рассмотрении данного вопроса, была проведена работа, результаты которой опубликованы в 1998 году в «Токайском журнале экспериментальной и клинической медицины» (The Tokai journal of Experimental and Clinical Medicine) в статье «Theronts of Ichthyophthirius multifiliis find their Fish Hosts with Complex Behavior Patterns and in Response to Different Chemical Signals». Авторы изучили особенности поисковых реакций инвазивных теронтов I. multifiliis во время представления им различных фракций и химических модификаций кожи рыб.
Свободноплавающие теронты проявили, по крайней мере, 6 различных поведенческих паттерна, 4 из которых стимулировались компонентами кожи рыб, то есть аминокислотами. Теронты перемещались по направлению к твердому субстрату (“останавливались и продолжали двигаться”) в ответ на высокомолекулярные гликоконъюгаты рыбий кожи, и они отклонялись (“останавливались и уходили”) при соприкосновении с другими высокомолекулярными гликоконъюгатами. После контакта с твердой поверхностью, они продолжали проникновение внутрь (“погружение при контакте”) в ответ на гликоконъюгаты рыбий кожи и двигались вдоль поверхности (“параллельно телу”), когда поверхность кожи содержала белки. Результаты показали, что одноклеточные паразиты рыбы, не имея нервной системы и клеточных хеморецепторов, могут проявлять разнообразные модели поискового поведения (по меньшей мере, 11) и способны отвечать на различные химические сигналы хозяина (по меньшей мере, 5).
Введение
Многие паразиты рыб заражают своего хозяина посредством активного проникновения на личиночной стадии. Этапы инвазии обычно происходят многократно, и возникает вопрос, должны ли паразиты определять хозяина для успешного переноса. Большое количество информации, касающейся обнаружения и идентификации хозяина, собрано для церкарий трематод (2-4). Данные многоклеточные организмы проявляют последовательный комплекс поведенческих моделей, когда приближаются и заражают хозяина, и в каждой из моделей они отвечают на независящие от хозяина сигналы, такие как течение воды, тень как стимул и, в особенности, различные химические сигналы. Эти изощренные меры идентификации хозяина играют ключевую роль в переносе паразита, независимо от количества создаваемых церкарий. В данном контексте интересно было пронаблюдать, каким образом лишенные нервной системы и клеточных рецепторов одноклеточные паразиты рыб находят и заражают своего хозяина. Авторы изучили способы обнаружения и идентификации хозяина теронтами одной из самых патогенных инфузорий — Ichthyophthirius multifiliis (1-7). Наиболее изученно поведение непаразитических инфузорий видов Парамециум и Тетрахимена. Они используют несколько форм движений, таких как плавание вперед и назад с различной скоростью, маневрирование по прямой, и данные модели перемещения, связаны с изменениями мембранного потенциала (8). Применяют ли теронты Ichthyophthirius multifiliis подобные простейшие модели перемещения, когда ищут хозяина?
Результаты
Авторы исследовали поведение теронтов I. multifiliis, когда они встречались с субстратом рыб. В качестве субстрата выступала живая, мертвая рыба, а также экстракт с поверхности кожи рыб и их изолированные и химически модифицированные смеси, интегрированные с агаром, либо пропущенные через мембранные фильтры. Теронты показали удивительное разнообразие ответов во время перемещения, в особенности, когда они приближались к субстрату и после того как они встречались с ним (Рисунок 1).
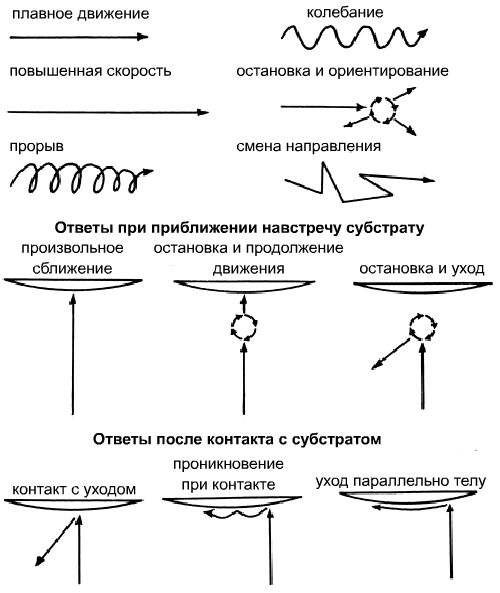
Рисунок 1. Тип плавательного поведения инвазивной стадии (теронт) паразитической ресничной инфузории I. multifiliis.
(1) Свободноплавающие теронты отвечали 4 моделями поведения по отношению к поверхности кожи рыб: ответ “прорыв” стимулировался неизвестными факторами в присутствии живой рыбы, но не мертвой, “колебание” на компоненты кожи рыб (не лягушки или улитки), Mr < и >1 кДа. Ответы “остановка и ориентирование” и “изменение направления” стимулировалось преимущественно аминокислотами с поверхности кожи рыб и неизвестными высокомолекулярными веществами кожи.
(2) Когда теронты приближались навстречу субстрату рыбы, они ориентировались в направлении хозяина, отвечая по схеме “остановка и продолжение движения” для высокомолекулярных гликопротеинов с поверхности кожи. Однако, заметно отклонялись в присутствии смесей поверхности кожи рыб: гликоконъюгаты поверхности кожи стимулировали реакцию остановка и уход”.
(3) После контакта с поверхностью субстрата теронты уже определяли хозяина. Они реагировали по типу “погружение при контакте” в присутствии определенных гликоконъюгаты типу “параллельно телу” в присутствии высокомолекулярных белков поверхности кожи рыб.
Обсуждение
Наблюдения Lom и Cercasovova (6) говорят о том, что теронты I. multifiliis не ориентируются по направлению к хозяину на сверхдальних дистанциях и концентрация теронтов около живой рыбы может быть даже ниже, чем в контрольных водных пробах (9). Церкарии, заражающие рыбу, также не способны к хемо-ориентации на дальние расстояния (2-4). Фактически, невозможно, чтобы стабильный химический концентрационный градиент устанавливался вокруг рыб на сверхдальних расстояниях, когда хозяин рассматривается как быстро перемещающийся. Тем не менее, церкарии трематод повышают возможность встречи с хозяином за счет ответа на сигналы в непосредственной близости от рыбы. Это справедливо и для теронтов I. multifiliis. В присутствии смесей поверхности кожи рыбок, свободноплавающие личинки отвечают, по крайней мере, 4 из 6 плавательных реакций. Модели перемещения могут значительно повысить шансы контакта с рядом находящейся рыбой. Во время движения навстречу субстрату организмы решают, будут они контактировать с объектом по типу “остановка и продолжение движения”, либо “остановка и уход”. После контакта с субстратом теронты отвечали двумя моделями (“погружение при контакте” и ”движение параллельно телу”), которые также имеют место быть у ресничных мирацидий трематод (6), сталкивающихся с хозяином улиткой, и, которые очевидно являются результатом идентификации хозяина.
Паразиты показали, по крайней мере, 11 различных моделей поведения и они отвечали, по меньшей мере, на 4 химических сигнала, испускаемых рыбой. Плавательное поведение теронтов I. multifiliis более разнообразное и более комплексное, чем у описываемых свободноживущих ресничных инфузорий. Результаты работы говорят о том, что активность инвазивной стадии паразита нуждается в комплексе механизмов поиска и идентификации хозяина, не зависящих от массовости воспроизводства.
——
по материалам: Haas Wilfried, Haberl Bernhard at al. Theronts of Ichthyophthirius multifiliis find their Fish Hosts with Complex Behavior Patterns and in Response to Different Chemical Signals. // Tokai J Exp Clin Med., Vol. 23, No. 6, pp.329-331, 1999
У кого фальш-дно и мощный водообмен ихтиофтириоз не страшен.
Для рыбовода эта стадия развития инфузории является ключом к лечению ихтиофтириоза. Если теронты не могут полагаться на хемотаксис и передвигаться на дальние дистанции, тогда высокий аквариум (скажем, цилиндрический) с решеткой по середине идеально подойдет для лечения болезни. Когда из цист на дне полезут теронты, они не смогут добраться до хозяина, и цикл заражения оборвется.